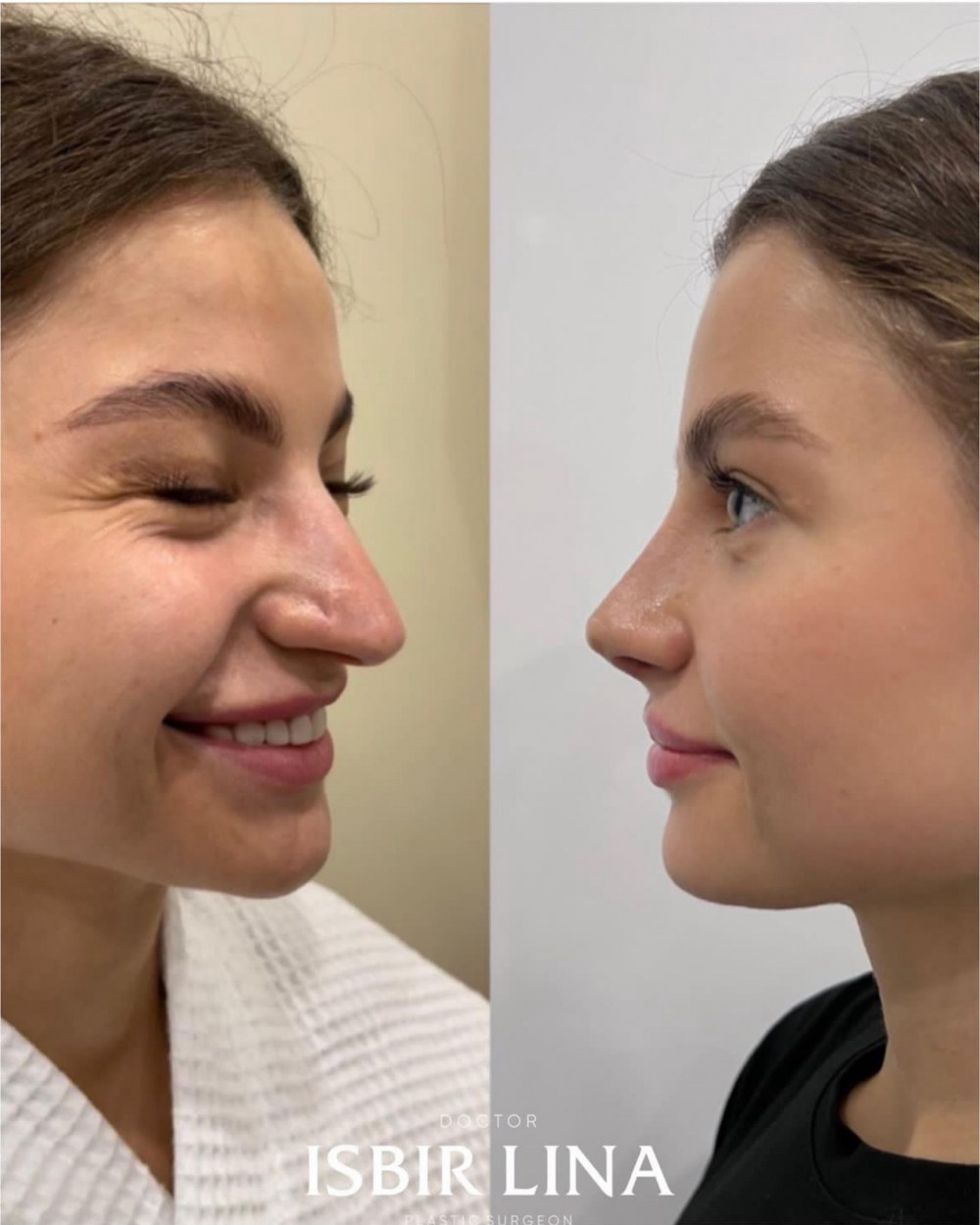
Как поддерживать здоровье и физическую активность во время дистанционного обучения Дети Как поддерживать здоровье и физическую активность во время дистанционного обучения